

Uncertainty Quantification for Deep Regression using Contextualised Normalizing Flows
Post hoc uncertainty quantification for deep regression using contextualized normalizing flows without retraining the base model.
adriel-sosa-marco
Computational biologist and data scientist with a track record of applying machine learning and statistical modelling to complex biological problems — from biomolecular structure prediction and de novo peptide design to metagenomic analysis and preclinical trial design.
My background combines rigorous quantitative training (Bayesian modelling, deep learning, MLOps) with hands-on experience across the full data pipeline — from experimental design through to deployment and reproducible reporting. I have worked at the interface of Biotech and AI in an industry research setting, contributing to work that reached commercial spinout stage and to a NeurIPS 2025 publication on uncertainty quantification in deep regression.
PhD
Lund University
MSc
University College Dublin
BSc (Hons)
University College Dublin
Post hoc uncertainty quantification for deep regression using contextualized normalizing flows without retraining the base model.
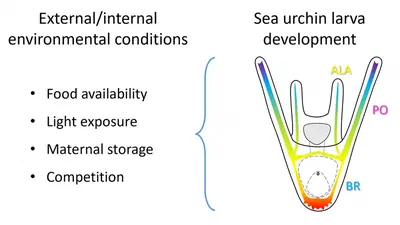
Phenotypic plasticity, the ability of a genotype to produce different phenotypes in response to environmental conditions, plays a crucial role in adaptation and evolution and can …
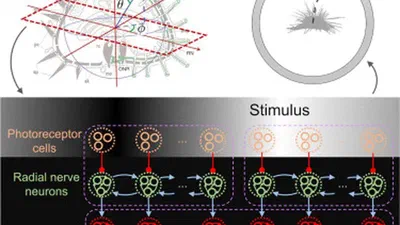
Sea urchins can detect light and move in relation to luminous stimuli despite lacking eyes. They presumably detect light through photoreceptor cells distributed on their body …